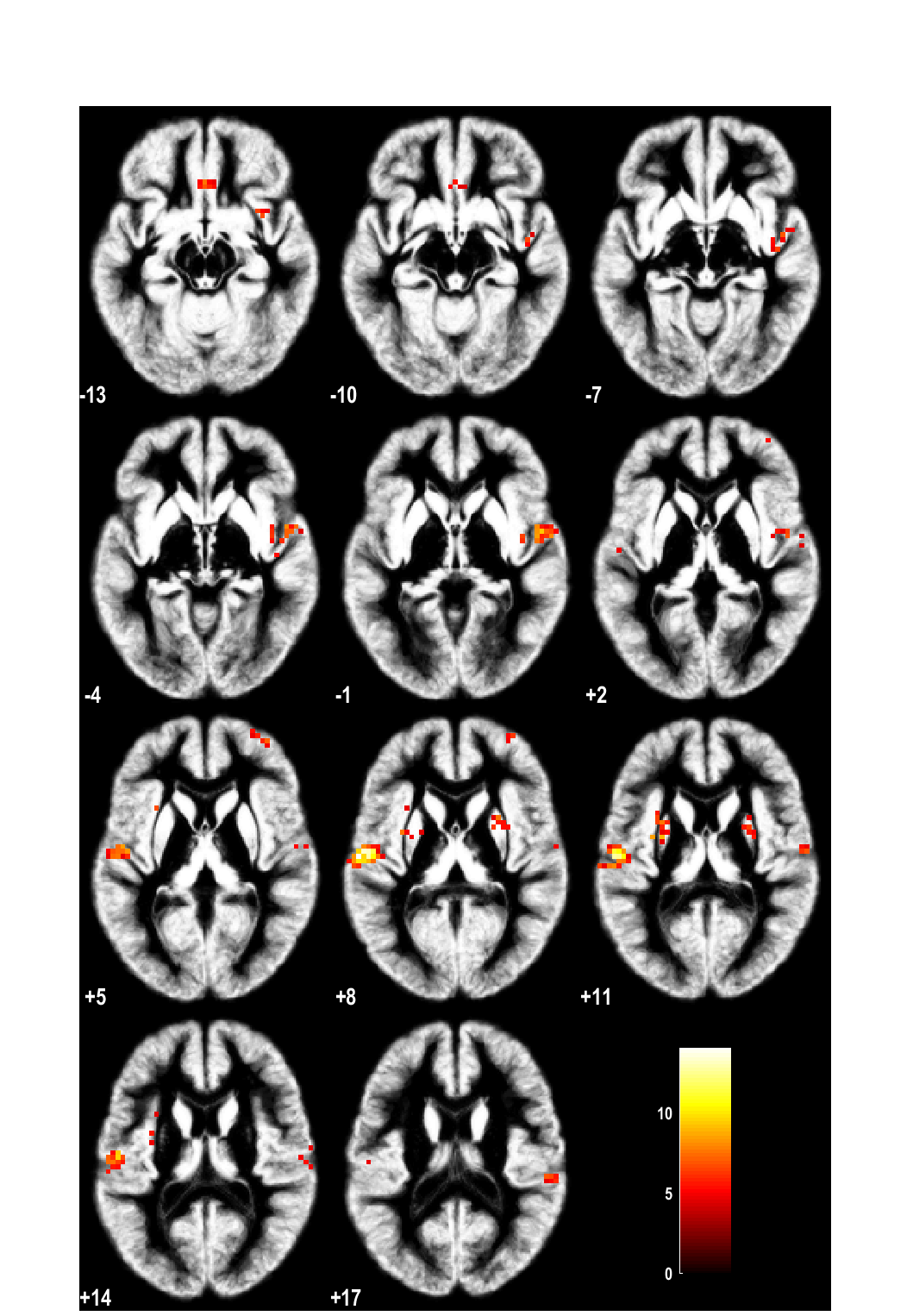
Brain activation after nasal histamine provocation in house dust mite allergic rhinitis patientsCallebaut I1, Steelant B1, Backaert W1, Peeters R2-3, Sunaert S2-3, Van Oudenhove L4-5*, Hellings PW1*1Allergy and Clinical Immunology Research Group, Department of Microbiology, Immunology and Transplantation, KU Leuven, Leuven, Belgium2Department of Imaging & Pathology, KU Leuven, Leuven, Belgium3Department of Radiology, University Hospitals Leuven, Leuven, Belgium4Laboratory for Brain-Gut Axis Studies (LaBGAS), Translational Research Center for Gastrointestinal Disorders (TARGID), Department of Chronic Diseases, Metabolism, and Ageing (CHROMETA), University of Leuven, Belgium5Cognitive and Affective Neuroscience Laboratory (CANlab), Center for Cognitive Neuroscience, Department of Psychological and Brain Sciences, Dartmouth College, Hanover, NH, USA*Joined senior authorshipTo the editor . The nasal mucosa is armed with a complex nervous system of sensory, sympathetic and parasympathetic nerves, allowing swift defensive responses to physical and chemical stimuli. In allergic rhinitis (AR) patients, nasal allergen deposition leads to mast cell activation with release of allergic mediators such as histamine. Apart from its direct effects on the surrounding tissue, histamine also activates sensory nerve endings giving rise to symptoms like sneezing, rhinorrhoea, and/or congestion(1). Activated nasal sensory nerves transmit action potentials to their cell bodies in the trigeminal ganglion and further to the midbrain where secondary synapses lead to the generation of central reflex signals. Despite activation of neural pathways in AR(2), it is not known which particular regions in the brain are activated by different nasal stimuli. Clinical studies using Positron Emission Tomography scans indicate that there is no isolated itch center in the brain but that different cortical centers are involved in the processing of itch(3, 4). Activation of the anterior cingulate cortex (ACC), the supplementary motor area (SMA), and the inferior paretial lobe partly explains the connection between itching and the related reflex of scratching(4). Using functional magnetic resonance imaging (fMRI), the activation of the superior temporal gyrus, insula and nucleus caudate following painful intranasal trigeminal stimulation has been shown(5). When asthmatic patients are challenged with metacholine or allergens, activity in ACC and insula was associated with markers of bronchial inflammation and obstruction(6).To fill the abovementioned knowledge gap, a prospective, single-blind, cross-over study was designed to investigate brain responses to nasal histamine provocation in healthy volunteers and AR patients.Eight house dust mite (HDM) AR patients and 7 non-allergic healthy controls (HC) were recruited at the outpatient clinic for Otorhinolaryngology of University Hospitals Leuven. HDM allergy was confirmed by a skin prick test. Relevant nasal anatomic abnormalities or rhinosinusitis were ruled out by nasal endoscopy. Non-allergic HC showed a negative skin prick test for all the tested allergens, showed no nasal symptoms and had normal nasal endoscopy. Patients of <18 and >50 years of age, having used nasal or oral steroid treatment <6weeks prior to the study or nasal or oral antihistamine treatment <4weeks prior to the study were excluded, as well as those with past or ongoing immunotherapy for HDM, asthma, smoking and clinical signs of rhinosinusitis or anatomic nasal deformities. Informed consent was signed by all participants. The study was approved by the local medical ethics committee of the University Hospitals Leuven (B322201215751).All HC and AR patients underwent a nasal provocation by means of a canulla placed under the nose with either nebulized sham solution (saline) or with histamine for 5 minutes while in supine position in the MR scanner on 2 separate days with a minimum of 1 week in between, and in a single-blinded and random order. An aerosol of 10 ml histamine HCl (16 mg/ml) or 10 ml saline was delivered via the canulla by means of air (8 bar) after 10 minutes of baseline scanning in a pharmacological (ph)MRI design. This concentration of histamine was chosen as optimal dose after a pilot study in 3 HCs, 1 birch and grass pollen AR patient and 1 HDM AR patient where the dose of histamine resulted in a reduction of 20% in the Peak Nasal Inspiratory Flow (PNIF). Moreover, patients did not had the urge to sneeze at this concentration, as was the case for the dose of 32 mg/ml.PNIF values were used for measuring nasal flow at baseline and after the nasal provocation at the end of the phMRI scan, as recommended(7). The best value out of three consecutive measurements with a variability of <10% was recorded. Changes in PNIF from baseline to post-provocation were compared between conditons (histamine & saline) as well as between groups (patients & controls) using marginal linear mixed models.phMRI data were preprocessed and analyzed as described previously(8, 9). The effect of interest for the present study was the group (patient versus controls)-by-substance (histamine versus saline)-by-time interaction effect, comparing the time-course of the brain response to histamine vs saline provocation between AR patients and controls. A whole-brain voxel-wise FWE-corrected threshold of p<0.05 was used combined with an extent threshold of k=10 voxels (corresponding to pFWE<0.001 at cluster level).In total, 8 HDM AR patients (5 females and 3 males) and 7 HC (5 females and 2 males) were recruited with a mean age of 22.5 ± 0.72 and 23.8 ± 1.11 years respectively. One female HDM AR and two female HC were excluded due to excessive head movement during MR scanning.After nasal provocation with saline, no significant decrease in PNIF was found compared to baseline in both groups (AR: 135 ± 61.82 l/min vs 137.5 ± 44.88 l/min, p=0.74; HC: 120 ± 36.74 vs 129 ± 31.30, p=0.46). Nasal provocation with histamine induced a significant decrease in PNIF in both HDM AR patients (158.8 ± 71.55 l/min vs 112.5 ± 83.67, p=0.0053) as well as in the HC (134.2 ± 27.64 l/min vs 85.83 ± 40.55, p=0.002).The analysis on PNIF values showed a significant condition-by-time (pre- to post-provocation) interaction effect (F(1,11)=28.8, p=0.0002), driven by a significant decrease in PNIF after histamine (-47.30±8.87, pHolm=0.0004), but not after saline (-5.81±5.96, pHolm=0.35) in the entire sample. No significant group-by-condition-by-time interaction effect was found (F(1,11)=0.09, p=0.78) indicating that the decrease from baseline after histamine compared to saline did not differ between patients and controls, with a significant decrease from baseline after histamine but not saline in both groups (p=0.002 and p=0.015, respectively).Brain regions showing a differential response to histamine versus saline in AR patients versus HCs included bilateral mid-/posterior insula, right anterior insula, bilateral postcentral/superior temporal gyrus/rolandic operculum (including secondary somatosensory cortex), bilateral putamen, left cerebellum (crus 1 & 2), right mid-occipital gyrus, bilateral medial orbital gyrus/gyrus rectus, and right middle/superior frontal gyrus (ventrolateral prefrontal cortex) (Table 1,Figure 1). Most of these differential responses were due to a stronger activation in controls vs AR patients, except for the right anterior insula, right middle occipital gyrus, right middle/superior frontal gyrus, and left cerebellum, where a stronger activation was observed in AR patients.