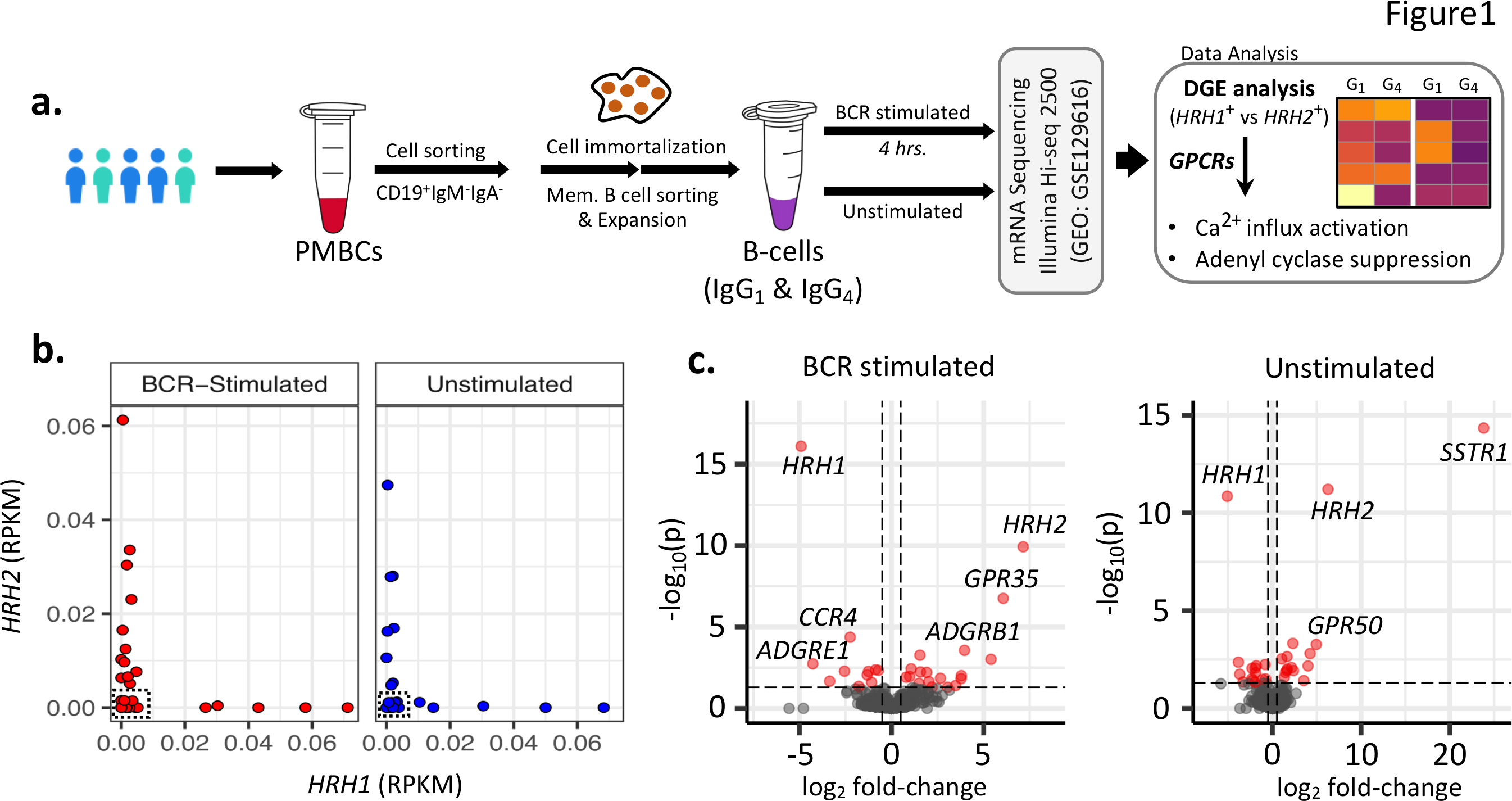
Title : Distinct and mutually exclusive Ca++ flux- and adenyl cyclase-inducing gene expression profiles of G-Protein-Coupled Receptors on human antigen-specific B cellsAuthors : Iris Chang1,2†, Abhinav Kaushik, PhD1,2†, Pattraporn Satitsuksanoa PhD1, Minglin Yang1, Laura Buergi Msc1, Stephan R. Schneider Msc1, Cezmi A. Akdis, MD1, Kari Nadeau MD, PhD2, Willem van de Veen, PhD1, Mübeccel Akdis, MD, PhD1*1 Swiss Institute of Allergy and Asthma Research (SIAF), University of Zürich, Davos, Switzerland.2 Sean N. Parker Center for Allergy and Asthma Research, Department of Medicine, Stanford University, Palo Alto, CA, USA.† Contributed equally* Corresponding authorB cells play an essential role in allergies by producing allergen-specific IgE, which is a prerequisite for allergen-induced degranulation of mast cells (MCs) and basophils. MCs, basophils, dendritic cells and bacteria are capable of releasing inflammatory mediators including histamine. Histamine is a bioactive amine that exerts its function through binding to histamine receptors (HRs), which are 7-transmembrane G-protein-coupled receptors (GPCRs). There are four types of HRs (HR1-4), wherein HR1 ligation triggers Ca2+ mobilization, HR2 stimulates and increases cAMP concentrations, and HR3 and HR4 inhibit cAMP accumulation1. In the presence of histamine in the environment, high affinity HR1is triggered causing cellular activation, followed by expression of 10 times lower affinity HR2 to regulate the over-inflammatory events. These HRs trigger different intracellular events upon activation, with HR1 as a Ca2+ flux-inducing activating receptor and HR2 as an adenyl cyclase-stimulating suppressive receptor 1,2. Therefore, to explore the response of B-cells in allergic diseases, we analyzed the expression profile of HRs and other GPCRs in B cell clones. We hypothesized that the expression profile of HRs (HR1+ vs HR2+ B cell clones) is associated with significant changes in the expression profile of other GPCRs that govern the downstream cascade of pathways associated with cAMP signaling or Ca2+ mobilization.A total of 27 IgG1 and IgG4 expressing B cell clones were isolated for gene expression analysis under BCR stimulated and unstimulated conditions (Figure 1A and Online Supplementary Methods) . Interestingly, we observed B-cell clones with mutually exclusive expression profile of HRH1 and HRH2 genes (Figure 1B), with more HRH1+ B-cell clones in BCR-stimulated samples than unstimulated samples. The subsequentHRH1+ vs HRH2+ differential gene expression analysis (Figure 1C ), reveal 27 differentially expressed (DE) GPCRs in unstimulated samples, with up-regulated P2RY13 and C5AR1genes in HRH2 + B-cell clones (Figure 2A) , which are associated with the cAMP signaling and suppressive pathway3,4. To further prioritize the DE GPCRs specifically associated with Ca2+ and cAMP signaling pathways, we reconstructed the co-expression networks and performed the weighted degree analysis across HRH1+ vs HRH2+ clones. The analysis reveals that the purinergic receptor family of GPCRs (i.e. P2RY1 , P2RY13 ) and complement component 5a receptor family of genes (i.e. C5AR1 and C5AR2 ) share highest degree of interactions. These genes are up-regulated inHRH2+ samples and are well-known to affect cAMP signaling pathway3,4 (Figure S1A ). Intriguingly, we also observed upregulation of GPR35 in HRH2 + B cells, which is associated in maintaining a low baseline Ca2+ level5. Similarly, we also observed up-regulation of GPR68 and GPR171 in HRH1 + B cells; both are known to stimulate Ca2+ flux (Online Supplementary Discussion) .Similarly, 28 GPCRs were differentially expressed in BCR-stimulated samples (Figure 2B ), including higher expression of serotonin receptor type 1A (HTR1A ) and HCAR1 (or GPR81 ) inHRH2+ samples, with a cAMP-linked suppressive function. In addition, we also observed upregulation of complement component 5a receptor family of genes (i.e., C5AR1 and C5AR2 ) and GPR35 , in agreement with the trend observed in unstimulatedHRH2 + B-cell clones. Surprisingly, we observed a higher expression of prostaglandin E2 receptor subtype EP4 (PTGER4) and adenosine A2A receptor (ADORA2A ) in HRH2+ samples3,6, which are known to be associated with activation of cAMP production and share the highest strength of interactions with the cAMP signaling sub-network (Figure S1B ). Among the up-regulated genes in HRH1 + samples, we found three Ca2+ mobilizing genes, i.e., GPR34 ,P2RY10 and PTAFR .The results reported in this study provides data for a novel hypothesis suggesting investigation of co-expressed genes that may play important synergistic or antagonistic regulatory roles in B-cell function.