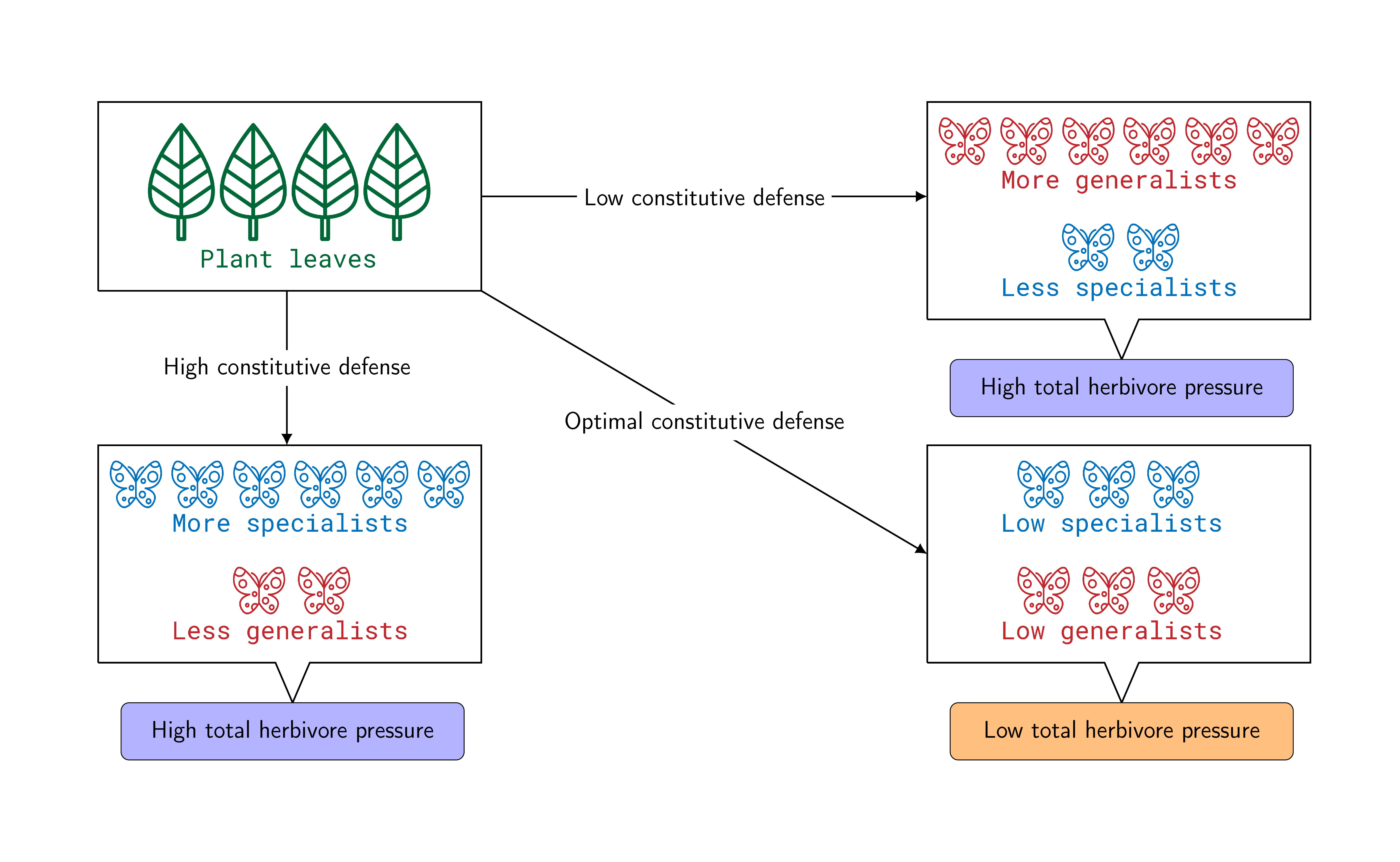
Brassicaceae plants have the glucosinolate-myrosinase defense system, jointly active against herbivory. Glucosinolates (GLS) are hydrolysed by myrosinase to produce isothiocyanates as soon as herbivory begins. Isothiocyanates exert detrimental effects on the feeding insects. However, constitutive GLS defense is observed to occur at levels that do not deter all insects from feeding. That prompts the question of why Brassicaceae plants have not evolved a high constitutive defense. The answer may lie in the contrasting relationship between plant defense and host plant preference of specialist and generalist herbivores. One of the reasons plants are in this dilemma is that they do not know what kind of herbivore will attack them in any given year, and thus have to be prepared for different possibilities. GLS content increases the susceptibility to specialist insects because these are attracted to plants with a high GLS content and are capable of coping with the toxin. In contrast, generalists are deterred by the plant GLS. Although GLS can attract the natural enemies (predators and parasitoids) of these herbivores, enemies can reduce herbivore pressure to some extent only. So, plants can be overrun by specialists if GLS content is too high, whereas generalists can invade the plants if it is too low. Therefore, an optimal constitutive plant defense can minimize the overall herbivore pressure. To explain optimal defense theoretically, we represent the contrasting host selection behavior of insect herbivores and, in addition, the emergence of their natural enemies by a non-autonomous ordinary differential equation model, where the independent variable is the plant GLS concentration. From the model, we quantify the optimal amount of GLS, which minimizes the total herbivore (specialists and generalists) pressure. That quite successfully explains the evolution of constitutive defense in plants from the perspective of optimality theory.