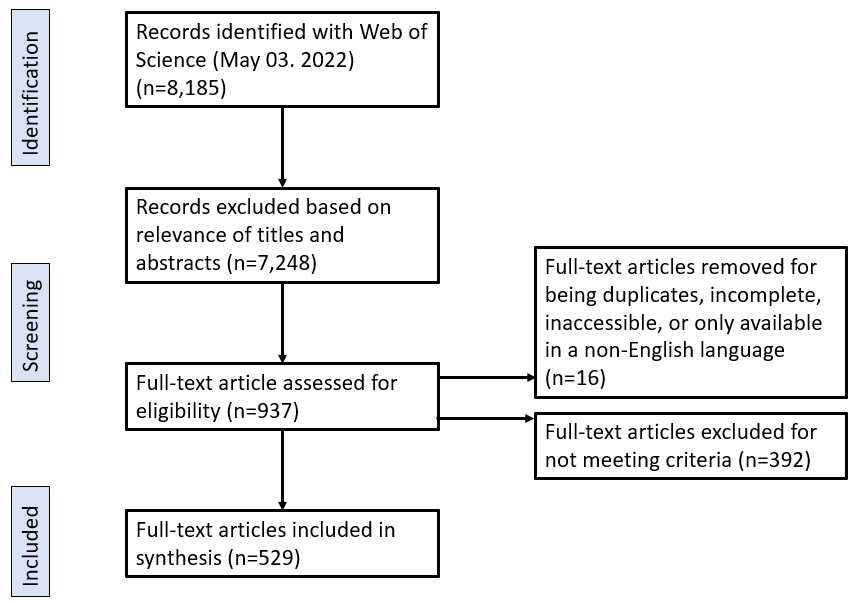
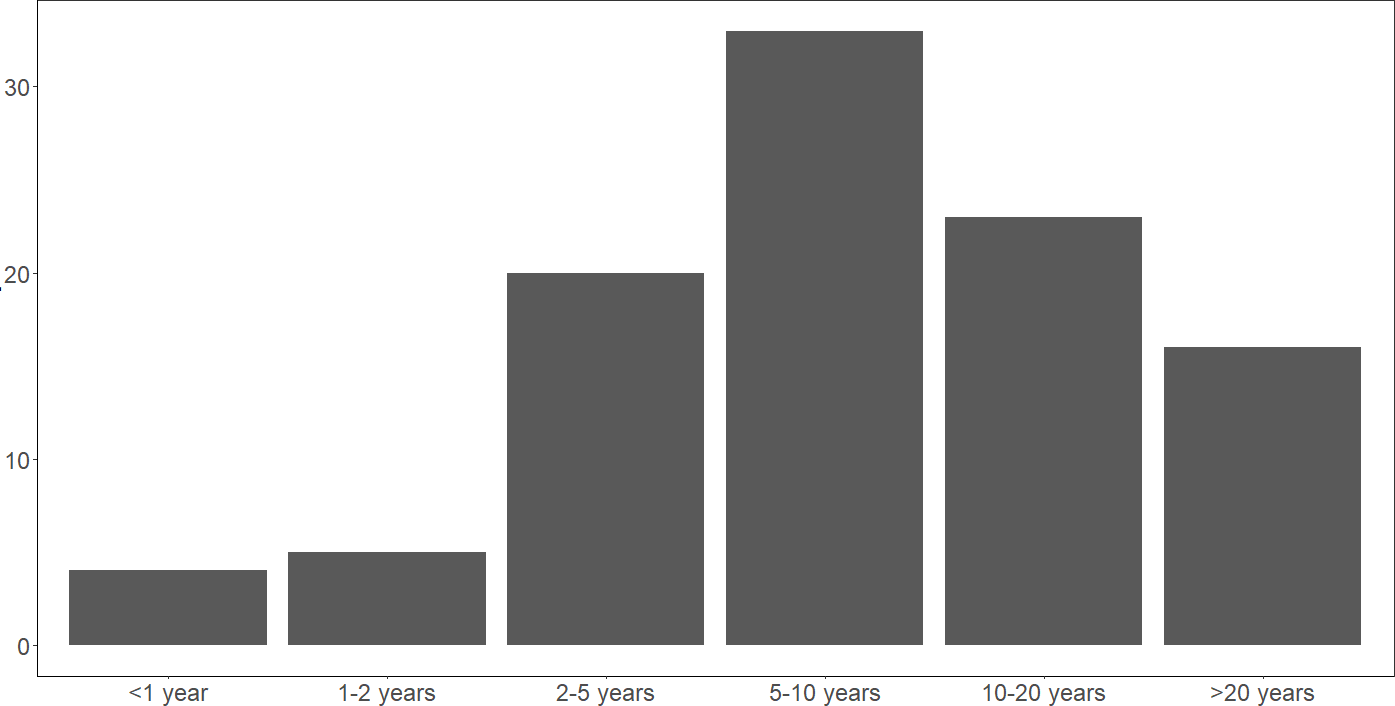Global patterns of reported human-wildlife interactions in areas of land-use changeReilly T. Jackson1, Tamika J. Lunn1, Nathaniel Mull1, Maureen R. McClung2, Kristian M. Forbes1Department of Biological Sciences, University of Arkansas, Fayetteville, Arkansas, USA 72701Department of Biology and Health Sciences, Hendrix College, Conway, Arkansas, USA 72032AbstractEmerging infectious diseases are one of the greatest and most pertinent threats to human health and security. Land-use changes, which are increasing globally, are a key driver of emerging infectious disease outbreaks; they increase contact between wildlife and humans and create opportunities for transmission of pathogens between them. While there is great emphasis to characterize the circumstances underlying disease outbreaks in search of mitigation strategies, a global synthesis of documented human-wildlife interactions in the context of land-use change has not been previously done. We conducted a scoping literature review to identify the geographic, taxonomic, and land-use change focus of reported interactions between wildlife, humans, and domestic animals, and discuss the implications of our results in the context of understanding high-risk settings for pathogen exposure. From 529 included articles, we show that human-wildlife interactions are most often reported in Asia and Africa and are most associated with agriculturalization on a global scale except in North America and Oceania, where urbanization was more commonly associated with interactions. Humans and domestic animals interacted with 1021 species of wildlife, including various amphibians, birds, mammals, and reptiles. Interactions with mammals were reported most often, mostly with species in the orders Artiodactyla, Carnivora, Primates, and Rodentia. The type of interactions, and thus potential for cross-species pathogen transmission, varied significantly among continent and wildlife taxa. Our review highlights increased human risks for wildlife pathogen exposure in areas of Africa and Asia with high rates of agriculturalization, and from even-toed ungulates, carnivores, primates, and rodents on a global scale. Further, we identified important gaps in knowledge, such as a lack of documented human interactions with wildlife in central and southwestern Asia and northern Africa and a surprising lack of documented human interactions with bats globally, despite their high number of synanthropic species and role as hosts of zoonoses.Keywords: agriculturalization, human-wildlife contact, land restoration, pathogen surveillance, spillover, urbanization, zoonosesRunning head: Human-wildlife contact under land-use changeContact author:Reilly T JacksonDepartment of Biological Sciences, University of Arkansas850 West Dickson Street, Fayetteville, USA 72701Email: [email protected], phone: +1 479-575-6701IntroductionEmerging infectious diseases (EIDs) are an increasing threat to global human health and security, as evidenced by the current monkeypox virus outbreak and COVID-19 pandemic (Daszak et al. 2000, Morens and Fauci 2013, Wang et al. 2022, Zumla et al. 2022). Most EIDs are zoonotic in origin, reaching human populations via transmission from wildlife and domestic animal host species (spillover), with the majority initiating in wildlife hosts (Jones et al. 2008). Once in human populations, zoonotic diseases have the potential to spread efficiently due to high population densities and connectedness in contemporary globalized societies, hindering the ease and potential efficacy of downstream mitigation efforts (Coltart et al. 2017).A major research priority is to understand the circumstances by which wildlife pathogens are transmitted to humans (Plowright and Hudson 2021). Exposure to pathogens at the human-wildlife interface is the obligatory first step in zoonotic spillover and is driven by multiple ecological mechanisms, including the distribution and abundance of reservoir host species, the prevalence and intensity of infection within reservoir hosts, and the persistence of pathogens once outside the host (Plowright et al. 2017, Wilkinson et al. 2018). Human exposure to wildlife pathogens can occur via direct (e.g., contact with wildlife through butchering and consumption) and indirect (e.g., contact with excreted pathogens in environments where human and wildlife activities overlap) mechanisms (Wolfe et al. 2005, Magouras et al. 2020). Additionally, domestic animals, including pets and livestock, can serve as intermediate (bridging) hosts between wildlife and humans, as occurred with several recent high profile disease outbreaks (e.g., pigs and horses for Nipah and Hendra viruses, respectively; Chua et al. 2000, Playford et al. 2010).One of the principal drivers of human exposure to wildlife pathogens is land-use change (LUC), or anthropogenically induced environmental changes (Foley et al. 2005, Woolhouse and Gowtage-Sequeria 2005, Jones et al. 2008, Gottdenker et al. 2014). Land-use change can impact the abundance and distribution of wildlife and shape wildlife-pathogen interactions, collectively increasing pathogen shedding by reservoir hosts and creating new contact opportunities that facilitate intra- and interspecies pathogen spread (Patz et al. 2004, Keesing et al. 2010, Jones et al. 2013, Faust et al. 2018, Mendoza et al. 2019, Carlson et al. 2022). For example, Nipah virus emergence in Malaysia is believed to have occurred due toPteropus bats moving to roost and feed in orchards surrounding pig farms following deforestation and El Niño-induced drought in their habitat (Chua et al. 2002); Nipah virus was shed in bat saliva and excreta, infecting pigs below, which in turn transmitted the virus to humans. Land-use change has also been associated with spillover ofBorrelia burgdorferi (the causative agent of Lyme disease), hantaviruses, ebolaviruses, and Hendra virus through effects on pathogen exposure (Allan et al. 2003, Wolfe et al. 2007, Plowright et al. 2011, Prist et al. 2017, Rulli et al. 2017).Existing review articles have evaluated links between LUC and pathogen spillover and emergence (Jones et al. 2013, Gottdenker et al. 2014, Johnson et al. 2015). These reviews focus on zoonotic disease outbreaks as a measure of spillover risk but do not evaluate exposure risk specifically. This nuance is crucial because documented disease outbreaks only capture a small fraction of total disease outbreaks (e.g., Glennon et al. 2019), and miss the many exposure opportunities that could have – but did not – lead to disease outbreaks (Plowright et al. 2017). Given that LUC primarily increase pathogen spillover and disease emergence in humans through effects on exposure risk, mechanistic insights into how LUC has influenced zoonotic spillover can be gained through evaluation of studies that link LUC with exposure. To this end, we conducted a scoping, quantitative literature review that characterizes the global breadth of studies that document human exposure to wildlife in the context of LUC to: 1) identify the geographic, taxonomic, and LUC focus of reported interactions; and 2) discuss and compare our results with previously identified geographic and taxonomic hotspots for spillover and emergence risk to highlight the most at risk settings and identify research needs.MethodsWe conducted a scoping literature search in Web of Science in May 2022 to identify empirical articles that report on wildlife interactions with humans and domestic animals in areas of LUC (Figure 1; a full description of the search strategy is available in the Supplementary Materials). To ensure that articles contained relevant information, we applied the following criteria. First, studies had to report human-wildlife interactions within the context of human-induced LUC that is occurring or has occurred in the study area and describe the type of modification. Second, studies had to identify the type of wildlife involved to at least order level. Third, studies had to report the type of human-wildlife interaction (direct, indirect, or domestic animal contact, see below). Fourth, studies must have been based on empirical data. We limited our scope to terrestrial and arboreal vertebrates since they are the overwhelming reservoir source of zoonotic disease outbreaks (Han et al. 2016, Olival et al. 2017).For each included article, we extracted the following information: the country where the interaction occurred, type of LUC, wildlife taxa involved, domestic animals involved, type of interaction, and standard journal article details (publication date, publishing journal). Land-use change was categorized into five types (Foley et al. 2005): (1) agriculturalization, (2) energy development, (3) land restoration, (4) resource extraction, and (5) urbanization (full definitions of each category are provided in the Supplementary Materials). Interactions between humans and wildlife were categorized into three types: (1) direct physical contact, such as humans touching or consuming wildlife and their effluent, (2) indirect contact, such as when humans and wildlife occupy the same areas but not necessarily simultaneously (e.g., humans observing wildlife on their property), or (3) domestic animal-mediated contact, as a way of quantifying the potential for exposure via intermediate host species that also have contact with humans (Table S1). Lastly, we collected information on the type of study (before-after comparison, cross-sectional, experimental, and longitudinal) and on techniques of data collection to understand methods applied within included studies.To investigate how the total number of publications reporting human-wildlife interactions varied by continent, taxa, interaction type, and LUC type, we used chi-square analyses. Post-hoc testing was done to understand interactions between every two categorical variable combination (package “chisq.posthoc.test”; Ebbert 2022). Due to a lack of studies on energy development and resource extraction, and given their similar characteristics with urbanization, these three LUC types were combined to permit more robust analyses. Due to lack of studies reporting human interactions with amphibians, we removed these records from analyses. For studies mentioning multiple continents, wildlife taxa, levels of interaction intensity, or LUC types, we counted the study for multiple categories (Gottdenker et al. 2014). Lastly, because of the differing zoonotic potential amongst wildlife orders, we used a chi-square analysis to compare publication count among orders within each class. Due to the large number of orders involved in interactions, we only included orders documented in 10 or more publications in this set of analyses.ResultsA total of 529 articles were identified that met our inclusion criteria (Figure 1; a full list of included articles and their citations are provided in the Supplementary Materials). Articles were published from 1994-2022 in 173 different journals and two pre-print servers. Over 85% of included articles were published since 2012 (n = 462), demonstrating a strong recent increase in relevant literature. Almost 70% of included articles included cross-sectional data (n = 357), followed by longitudinal (n = 231), experimental (n = 15), and before-after comparisons (n = 8). There were 13 main types of data collection techniques, with the most common methods including human interviews (n = 298), structured observation (n = 137), and analysis of government, non-governmental organization, or public records (n = 121; Table S2).We identified differences in reporting of human-wildlife interactions within all four categories. Human-wildlife interactions were reported in 96 countries, including all continents except Antarctica, with most reported in Asia and Africa and the least reported in Europe and Oceania (χ2 = 246.27, df = 5, P < 0.001; Figure 2). Agriculturalization was the most common LUC type reported (n = 407), followed by urbanization (n = 285), and restoration (χ2 = 37.805, df = 2, P < 0.001; n = 263). Human and domestic animals interacted with wildlife belonging to 50 distinct orders and 1,021 species, all of which fall into amphibians, birds, mammals, and reptiles (Classes: Amphibia, Aves, Mammalia, and Reptilia, respectively). Human interactions with mammals were reported most often (n = 493), followed by birds (n = 74) and reptiles (χ2 = 616.19, df = 2, P < 0.001; n = 45). Wildlife was documented to interact with 21 different species of domestic animal, with cows (n = 99), goats (n = 71), and sheep (n = 63) most reported. Indirect contacts (i.e., human spatial proximity to wildlife without physical contact) were the most common type of human-wildlife interaction (n = 400), followed by domestic animal-mediated contact (i.e., interactions between wildlife and domestic animals; n = 223), and direct contact (i.e., physical contact between a human and wildlife or their effluent; χ2 = 76.14, df = 2, P < 0.001; n = 219).We identified interactions in four of the six possible pair-wise variable combinations. We detected an interactive effect between wildlife taxa and interaction type (χ2 = 11.281, df = 4, P = 0.023). Interaction type did not vary within birds or mammals; however, within reptiles, direct and domestic animal-mediated interactions were significantly more common than indirect interactions (P = 0.029). There was an interactive effect between LUC type and continent (χ2 = 34.672, df = 10, P < 0.001), with reported interactions in areas of urbanization in Oceania and North America more common than in other LUC types on these continents (P ≥ 0.001; Figures 3 and 4). We detected an interactive effect between wildlife taxa and continent (χ2 = 18.456, df = 10, P = 0.047), with reported interactions with mammals in Asia more common than reports of interactions with other taxa (P = 0.028). Lastly, there was an interactive effect between continent and interaction type (χ2 = 19.504, df = 10, P = 0.034), with reports of direct interactions in Europe less common than other interaction types (P = 0.038; Figure 5). There were no interactive effects between LUC type and wildlife taxa (χ2 = 3.4819, df = 4, P = 0.481) or LUC type and interaction type (χ2 = 6.733, df = 4, P = 0.151).Order diversity within taxa varied considerably. Humans and domestic animals interacted with one amphibian, 27 avian, 19 mammalian, and three reptilian orders (Figure 6). Our analysis of the number of publications reporting interactions with different taxa varied among avian (χ2 = 14.44, df = 6, P = 0.025), mammalian (χ2 = 829.45, df = 10, P < 0.001) and reptilian (χ2 = 17.10, df = 2, P < 0.001) orders. Across avian orders, only Galliformes were reported more often than expected, with all other orders (Accipitriformes, Anseriformes, Charadriiformes, Columbiformes, Passeriformes, and Psittaciformes) reported as often as expected (P = 0.025). Within Mammalia, the orders Artiodactyla, Carnivora, Primates, and Rodentia were reported more often in the literature than expected; the orders Chiroptera, Cingulata, Didelphimorphia, Lagomorpha, Perissodactyla, and Pholidota were reported less often than expected; and the order Proboscidea was reported as often as expected (P < 0.001). Within Reptilia, the order Crocodilia was reported more often than expected, the order Testudines was reported less often than expected, and the order Squamata was reported as expected (P = 0.002).DiscussionThis study is the first to characterize publication trends documenting human and domestic animal exposure to wildlife in the context of LUC. We show that human-wildlife interactions are most often reported in Asia and Africa and are most commonly associated with agriculturalization on a global scale but with urbanization in North America and Oceania, specifically. Humans and domestic animals interacted with over 1000 species of wildlife, but interactions with mammals were documented most often, particularly with members of the Artiodactyla (even-toed ungulates), Carnivora, Primate, and Rodentia orders. Interaction type varied among continent and wildlife taxa, which has important implications for the risk of zoonotic pathogen spillover following wildlife exposure. We further focus on understanding how these geographic and taxonomic hotspots of human-wildlife interactions relate to known zoonotic disease emergence and identify areas for future research that will facilitate a comprehensive picture of zoonotic disease risk.