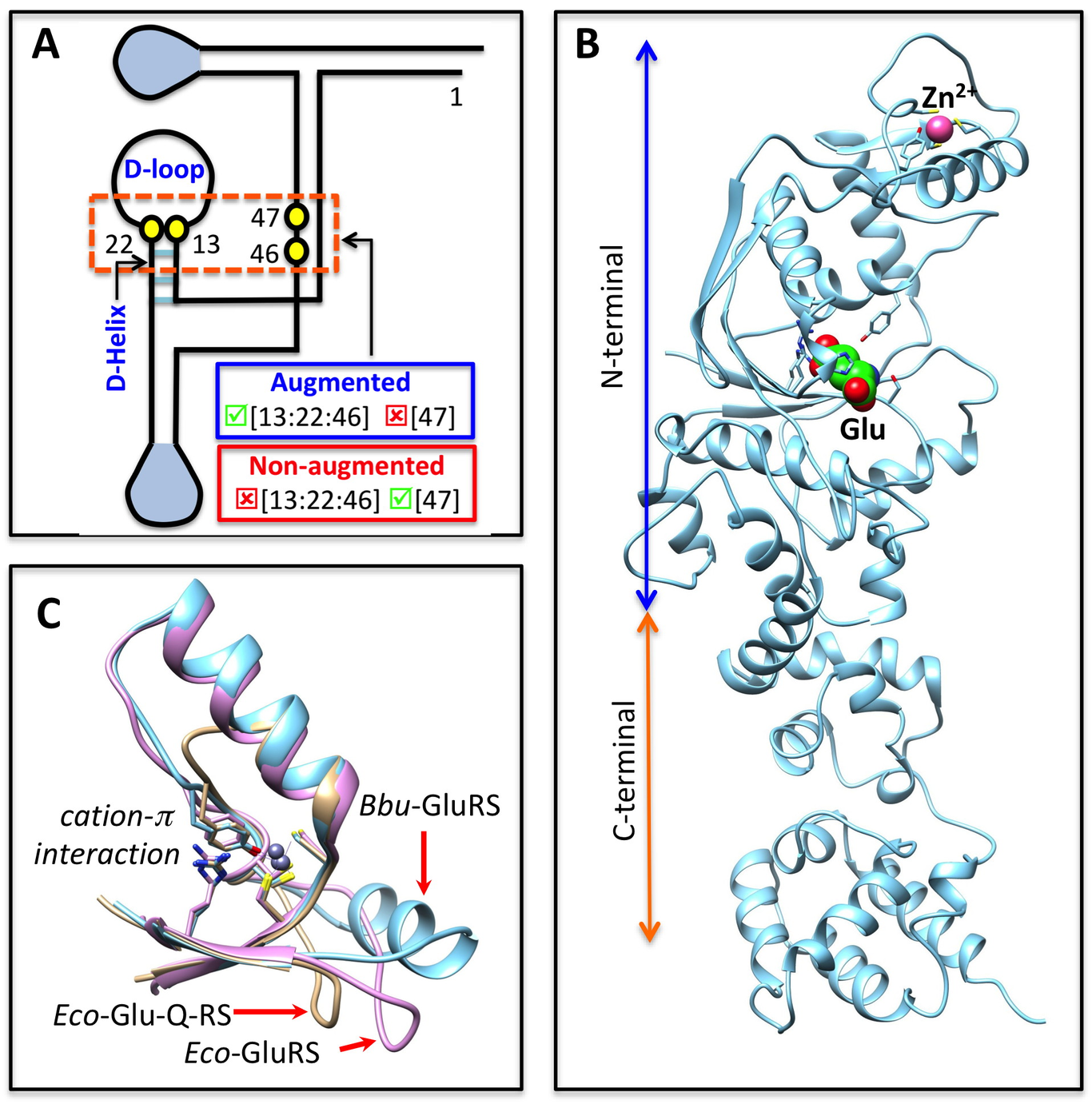
The canonical function of glutamyl-tRNA synthetase (GluRS) is to glutamylate tRNA Glu. Yet, not all bacterial GluRSs glutamylate tRNA Glu; many glutamylate both tRNA Glu and tRNA Gln, while some glutamylate only tRNA Gln and not the cognate substrate tRNA Glu. Understanding the basis of this unique tRNA Glx-specificity is important. Mutational studies have hinted at hotspot residues, both on tRNA Glx and GluRS, that play crucial roles in tRNA Glx-specificity. But the underlying structural basis remains unexplored. Majority of biochemical studies related to tRNA Glx-specificity have been performed on GluRS from Escherichia coli and other proteobacterial species. However, since the early crystal structures of GluRS and tRNA Glu-bound GluRS were from non-proteobacterial species ( Thermus thermophilus), the proteobacterial biochemical data have often been interpreted in the context of non-proteobacterial GluRS structures. Marked differences between proteo- and non-proteobacterial GluRSs have been demonstrated and therefore it is important that tRNA Glx-specificity be understood vis-a-vis proteobacterial GluRS structures. Towards this goal we have solved the crystal structure of GluRS from E. coli. Using the solved structure and several other currently available proteo- and non-proteobacterial GluRS crystal structures, we have probed the structural basis of tRNA Glx-specificity of bacterial GluRSs. Specifically, our analysis suggests a unique role played by a tRNA Glx D-helix contacting loop of GluRS in modulation of tRNA Gln-specificity. While earlier studies had identified functional hotspots on tRNA Glx that controlled tRNA Glx-specificity of GluRS, this is the first report of complementary signatures of tRNA Glx-specificity in GluRS.